Centipedes or chilopods are part of a larger group of arthropods called the Myriapoda.
There are five extant centipede orders: Scutigeromorpha, Lithobiomorpha, Scolopendromorpha, Geophilomorpha and Craterostigmomorpha.
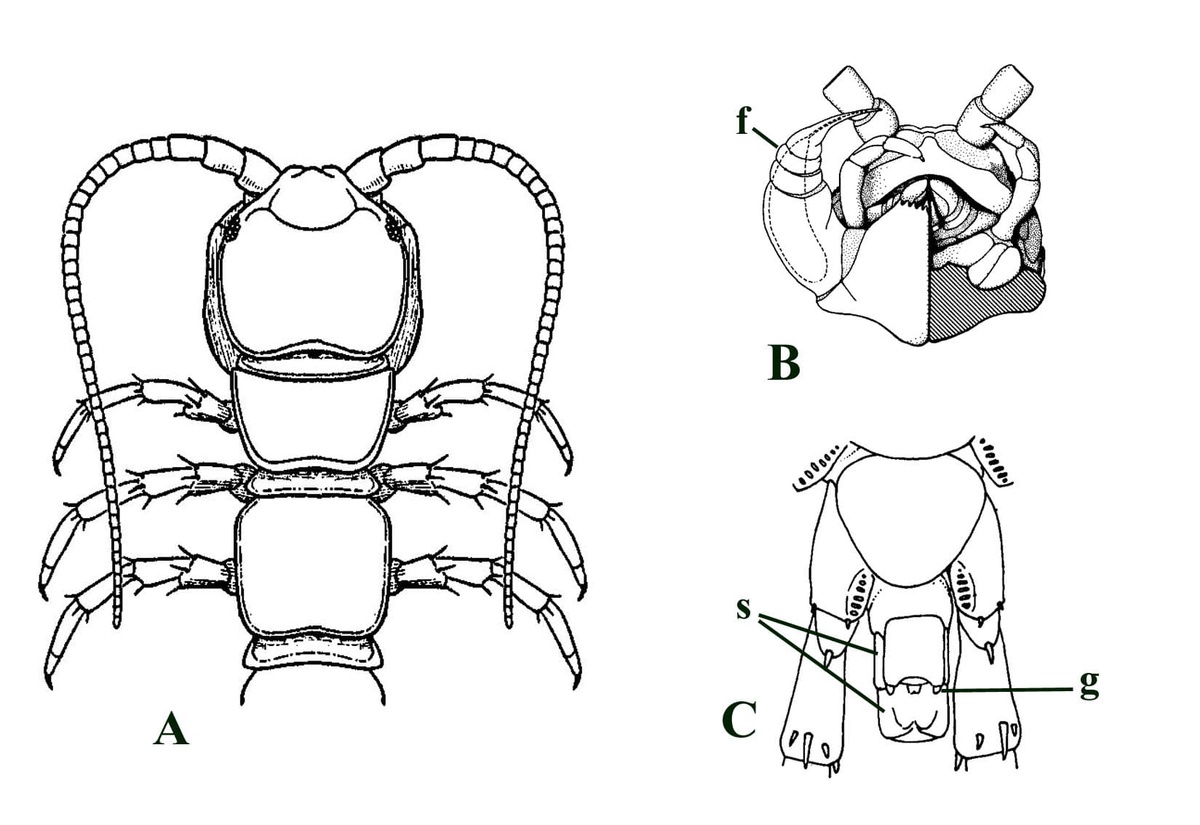
Centipedes are 'lung' or trachea-bearing terrestrial myriapods with a distinct head and a segmented elongate and flattened worm-like body with fifteen or more pairs of walking legs. The walking legs are evenly spaced with one pair per body segment. The cuticle of centipedes is non-mineralised and relatively permeable. Characteristically centipedes have their genital appendages on the last body segment and the appendages of the first segment behind the head capsule are developed into forcipules or "poison claws" (see inset below).
- Fossil Record
-
On the whole, as with other myriapods, the centipede fossil record is very poor. This is due mainly to the poor preservation potential of the non-mineralised exoskeleton within a terrestrial environment and also the fact that modern centipedes tend to eat their old cuticle after it has been shed during moulting. Therefore in sedimentary rocks where these fossils are found there must have been special circumstances at the time of deposition to allow preservation (see also the section on Taphonomy).
Centipedes range from the Late Silurian to the present day. The earliest known centipedes are recorded from macerates taken from the Upper Silurian Ludlow Bone Bed in Shropshire, UK (Jeram et al. 1990). Specimens have been recorded from the Cenozoic, primarily the Baltic and Dominican ambers, though only a few are known from the Mesozoic (e.g. Wilson 2001). Palaeozoic chilopods have been described from the Upper Pennsylvanian Mazon Creek fauna (Mundel 1979) and have also been recorded from the Upper Carboniferous of Canada (Matthew 1894) and France (Almond 1985). Centipedes have also been described from Middle Devonian of Gilboa, New York (Shear & Bonamo 1988), and of course the Early Devonian Rhynie chert (e.g. Shear et al. 1998, Anderson & Trewin 2003). The one unequivocal centipede described to date from the Rhynie and Windyfield cherts is the scutigeromorph Crussolum sp. (Anderson & Trewin 2003).
New chilopod material recently discovered in the Windyfield chert is currently being described and when published will be added to this resource page in due course.
- Morphology
-
Only fragmentary remains of Crussolum sp. have been found and hence the morphology of the entire animal remains unknown. However, the material we have suggests it had close affinities with modern scutigeromorph centipedes which are believed to represent the most primitive of the extant centipede orders. Extrapolation using the dimensions of the known elements, shown below, suggests the animal attained a length of approximately 35mm (Anderson & Trewin 2003).
Head
Of the head of Crussolum sp. only a few, but diagnostic fragmentary elements have been found. These include remains of the forcipular segment (the segment bearing the 'poison claws'), an antenna and probable mouthparts. For the purposes of this site we will concentrate on the forcipules and the antennae.
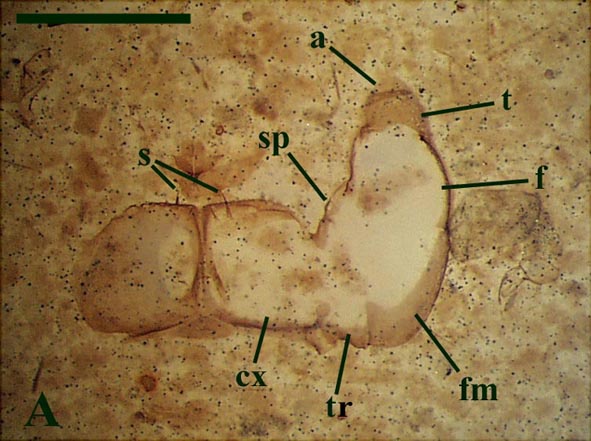
The forcipular segment (see inset below left) of Crussolum is closely comparable to that of modern scutigeromorph centipedes (see inset below right). The sternum and coxae are fused to form two coxosternal plates (coxosternae), split by a medial suture. The anterior margin of these plates bear a series of socketed anteriorly directed spines. A forcipular appendage comprises a very short 'trochanter' followed by a stout, robust femuroid. The femuroid article, like that in modern scutigeromorphs, bears a single socketed spine on the anterior surface which in life probably helped in pinioning prey items. Following the femuroid are three more articles; the femur, the tibia and finally the characteristic apical claw.

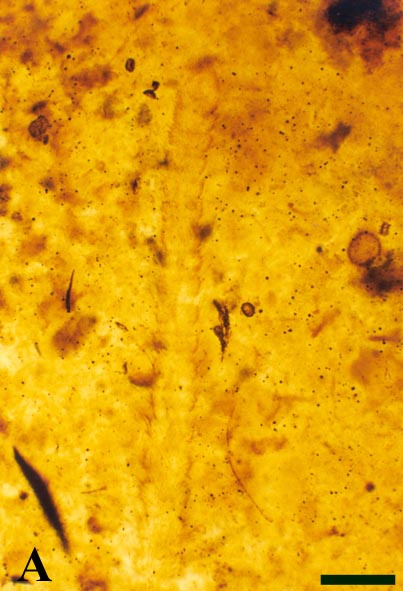
A distal end of a single antenna of Crussolum has been found (see inset below left). It is 2.5mm long and comprises at least 21 individual articles (segments). Each article is covered by dense anteriorly directed hairs or setae. The most distal or apical article appears swollen and bears three short, stout projections. Although not a complete antennal series, this again is quite comparable with antennae seen in modern scutigeromorph centipedes (see inset below right).

Body and Appendages
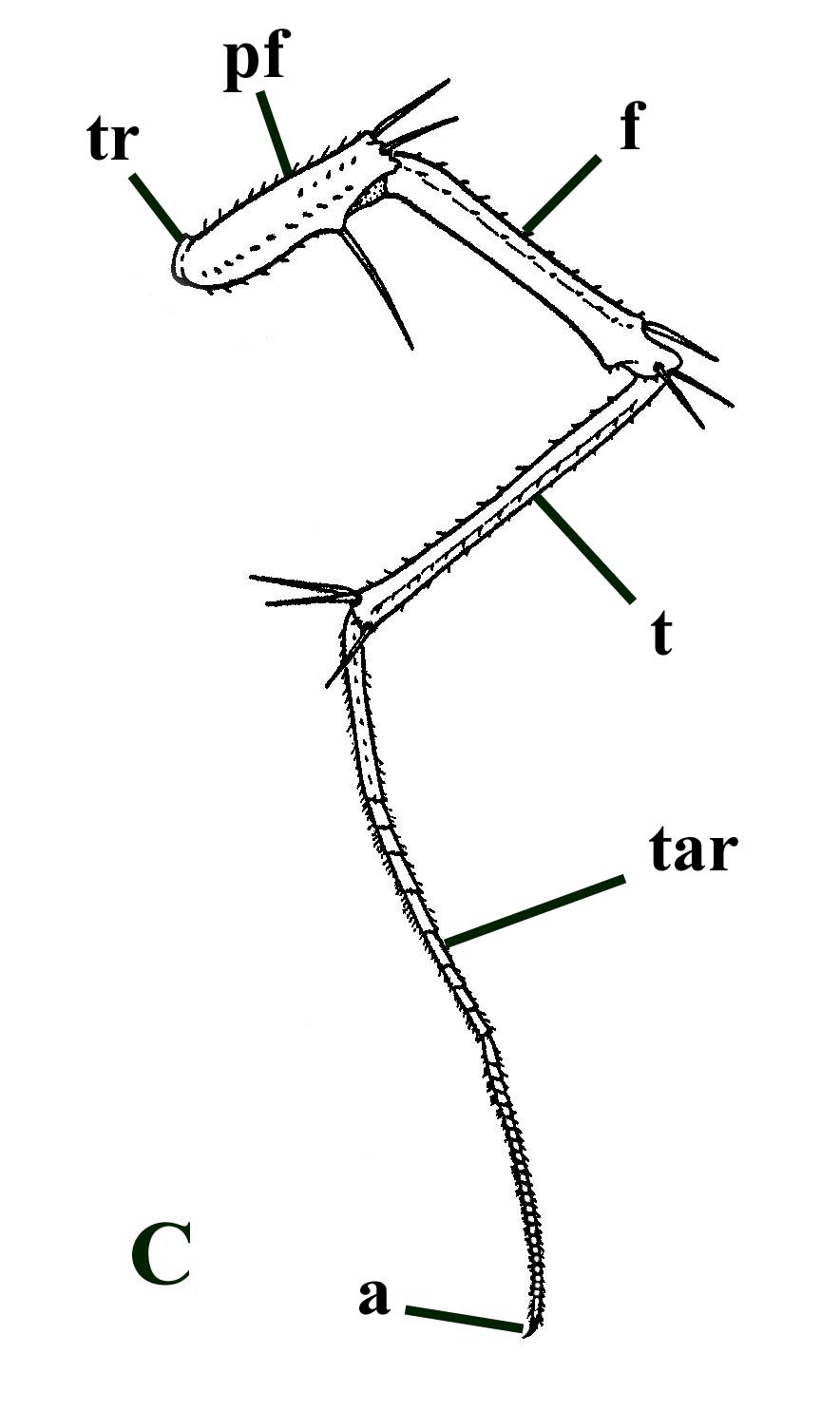
As far as the body or trunk segments are concerned only a few arch-shaped cross-sections through featureless cuticle sheets have been found associated with the material. However, the appendages are better represented, primarily comprising disarticulated podomeres. These exhibit a characteristic ornament very similar to modern scutigeromorph centipedes (see insets below). The podomeres, particularly the prefemur, femur and tibia exhibit longitudinal serrate or 'saw blade' ridges of distally pointing spines. There are typically five discrete ridges giving these podomeres a pentagonal outline in transverse cross-section (see inset below, top left). The distal ends of at least the femur and tibia of Crussolum bear distinctive socketed spines (see inset below, bottom left).


- Palaeoecology
-
Since centipedes possess a relatively permeable cuticle, and to prevent their bodies from drying out, they require damp environments in which to live; for example in soil, within plant litter, and under stones.
They are very agile animals and are voracious predators, particularly, but not exclusively, of other small invertebrates. There is no reason to suppose centipedes in the Early Devonian were really any different to their modern counterparts.
Crussolum was probably one of the most active predators in the Rhynie palaeoecosystem, perhaps hiding within plant litter or under degraded fragments of sinter and ambushing any potential prey item that strayed too close.