In 2000, field surveys conducted by the University of Aberdeen resulted in the discovery of several chert float blocks near Castle Hill farm approximately 1.5km east of the known Rhynie and Windyfield chert sites (see Rice et al. 2002). The cutting and preparation of one of these chert blocks led to the discovery of a new branchiopod crustacean, named Castracollis wilsonae and recently described by Fayers and Trewin (2003). Numerous specimens were discovered in one 50mm thick interval of the chert block, comprising near complete and fragmentary remains. This is the first new crustacean described from the chert since Lepidocaris rhyniensis (Scourfield 1926, 1940c).
- Relationships
-
Comprising a long multi-segmented thorax and abdomen, the thorax bearing two distinct sets of numerous appendages; and possibly possessing a large cephalothoracic shield, this animal belongs to the phyllopod lineage of the Branchiopoda and appears to be closely related to the Calmanostraca (a phyllopod group that includes the modern Tadpole Shrimps (the Notostraca) and the extinct Kazacharthra). It also shows some similarities in terms of its head morphology with the Onychura (the other phyllopod group that includes the Water Fleas or Cladocera and the Clam Shrimps or Conchostraca). For reviews of these other branchiopods and their relationships refer to Walossek (1993, 1995).
Castracollis is the earliest known calmanostracan-like branchiopod in the fossil record. Although track-ways attributed to similar animals are known from the Lower Devonian, until now the earliest known fossil Calmanostraca have been recorded from the Carboniferous.
The following table shows some of the differences and similarities between the morphologies of Castracollis, the Notostraca, Kazacharthra, Lipostraca and Conchostraca:
Castracollis Notostracans (tadpole shrimps)
Kazacharthrans Lipostracans e.g.: Lepidocaris
Conchostracans (clam shrimps)
Possible cephalothoracic shield covering head and thorax Single, broad cephalothoracic shield covering head and thorax Single, broad, often ornamented, cephalothoracic shield covering head and thorax No head shield in adults Bivalved cephalothoracic shield enclosing body Long biramous 2nd antennae Reduced uniramous 2nd antennae in adults Reduced biramous 2nd antennae Long biramous 2nd antennae Long biramous 2nd antennae Distinct anterior and posterior series of trunk limbs: 21 - 26 pairs of which the posterior 10 -15 are relatively small (and possibly post genital) Distinct anterior and posterior series of trunk limbs: 35 - 71 pairs of which the posterior 29 - 52 are relatively small and post genital with variable number of leg pairs per body segment 11 pairs of trunk limbs (no distinct posterior series) 11 pairs of trunk limbs, of which the first pair are modified maxillae. 10 -32 pairs of trunk limbs; 1 - 16 of which are post genital Lateral scales on leg-bearing body segments Lateral scales absent Lateral scales absent Lateral scales on leg-bearing body segments Lateral scales absent - Morphology
-
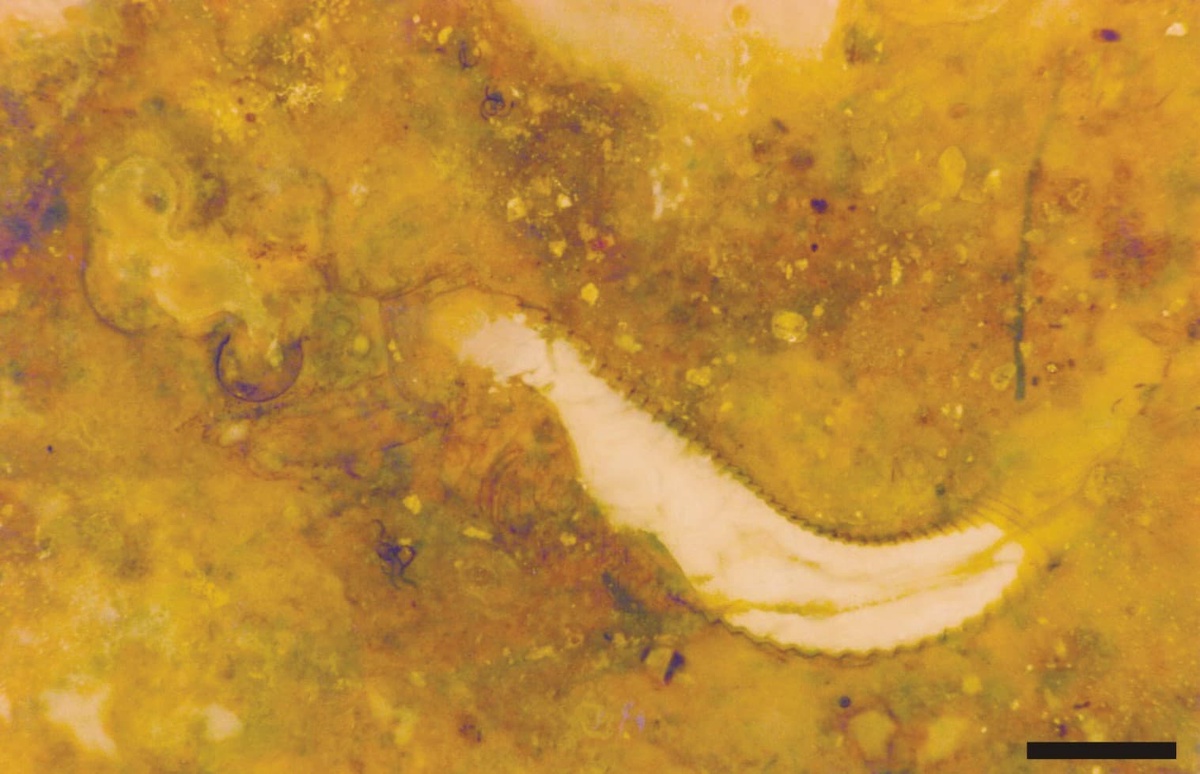
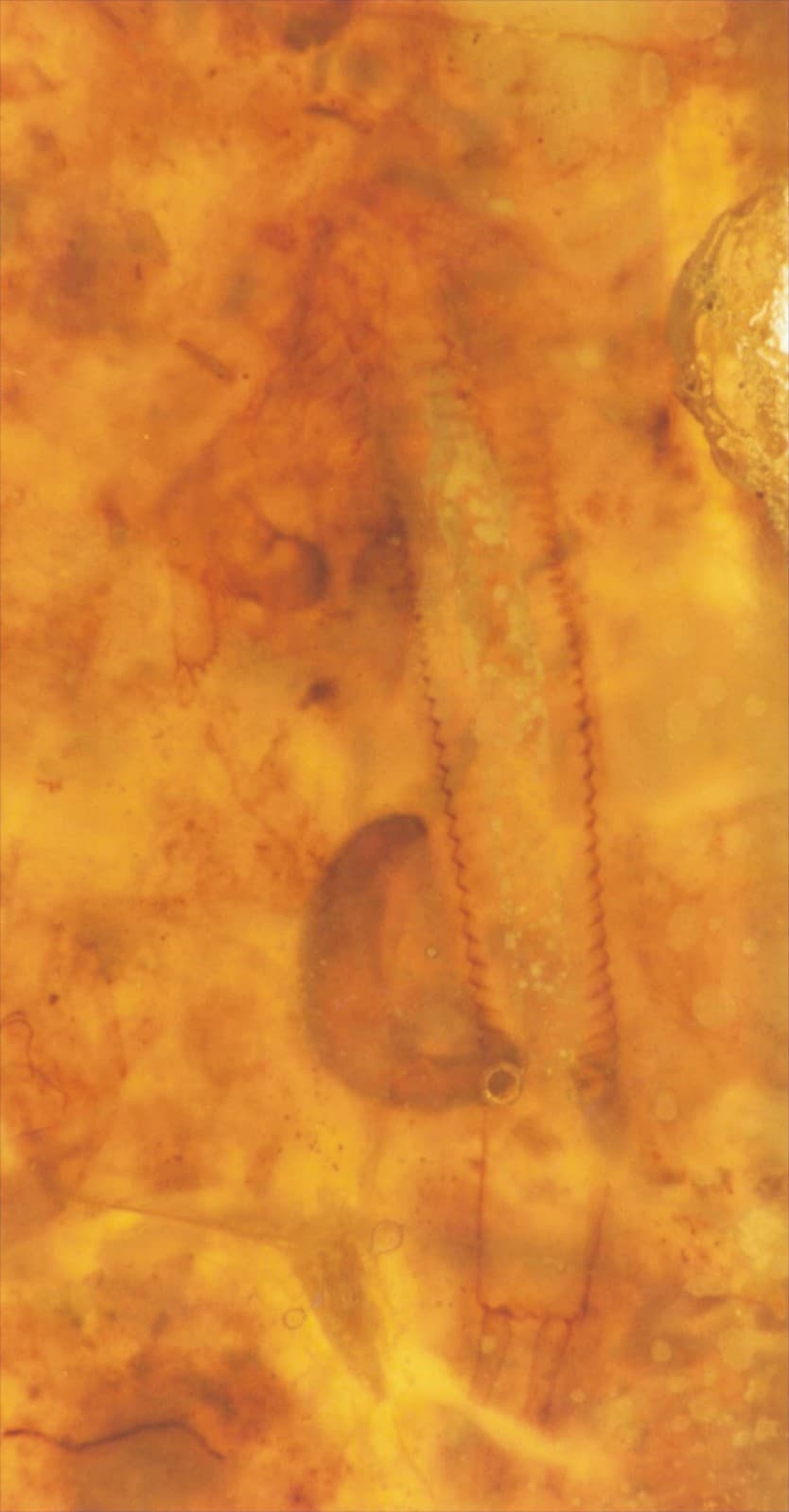
Castracollis was a small branchiopod, the largest specimens found being 8mm in length. The majority of the specimens are fragmented, and the more complete specimens show damage to the anterior of the trunk and the head is often detached. This suggests most are exuviae or the moults of the animal.
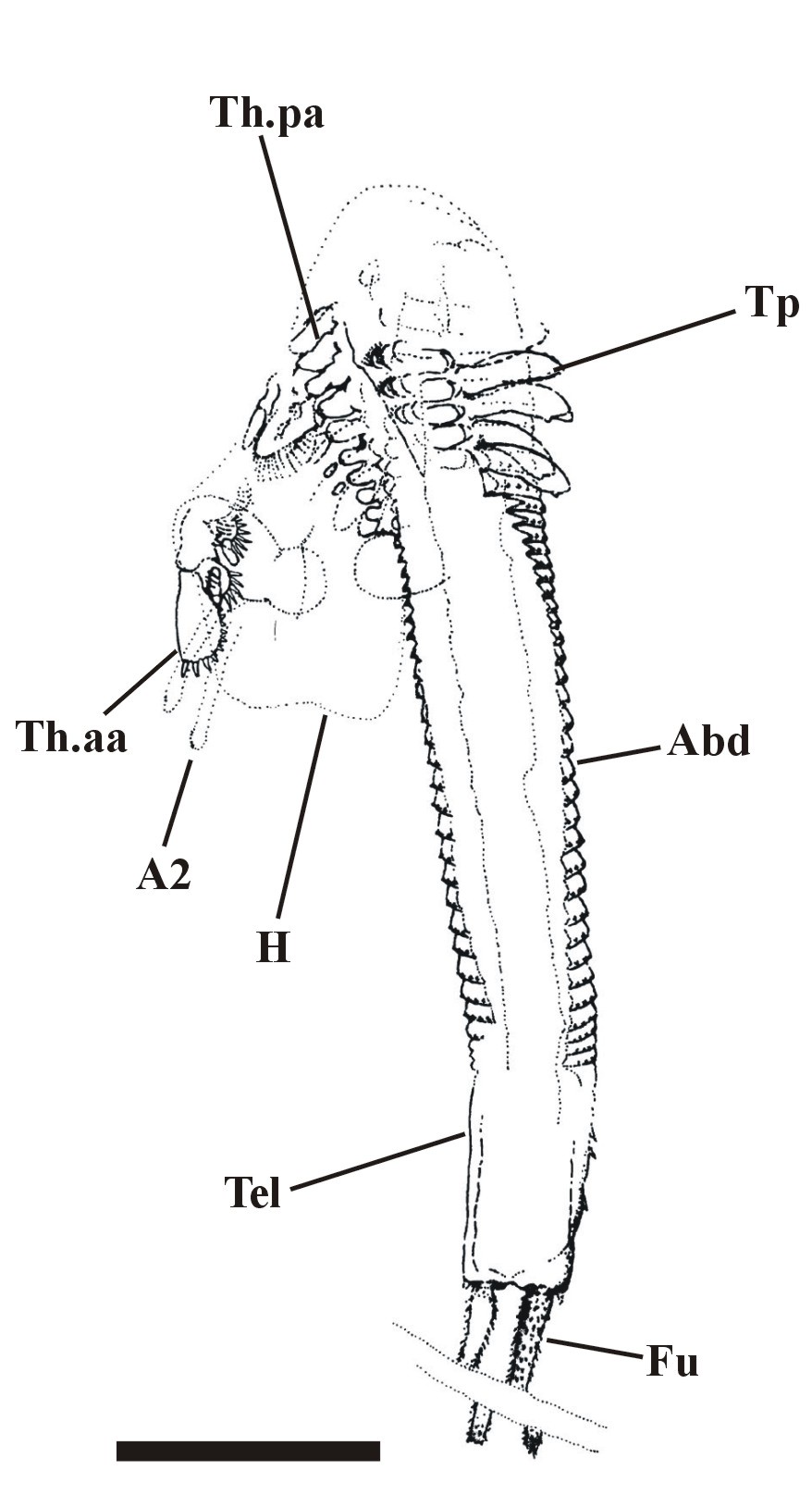
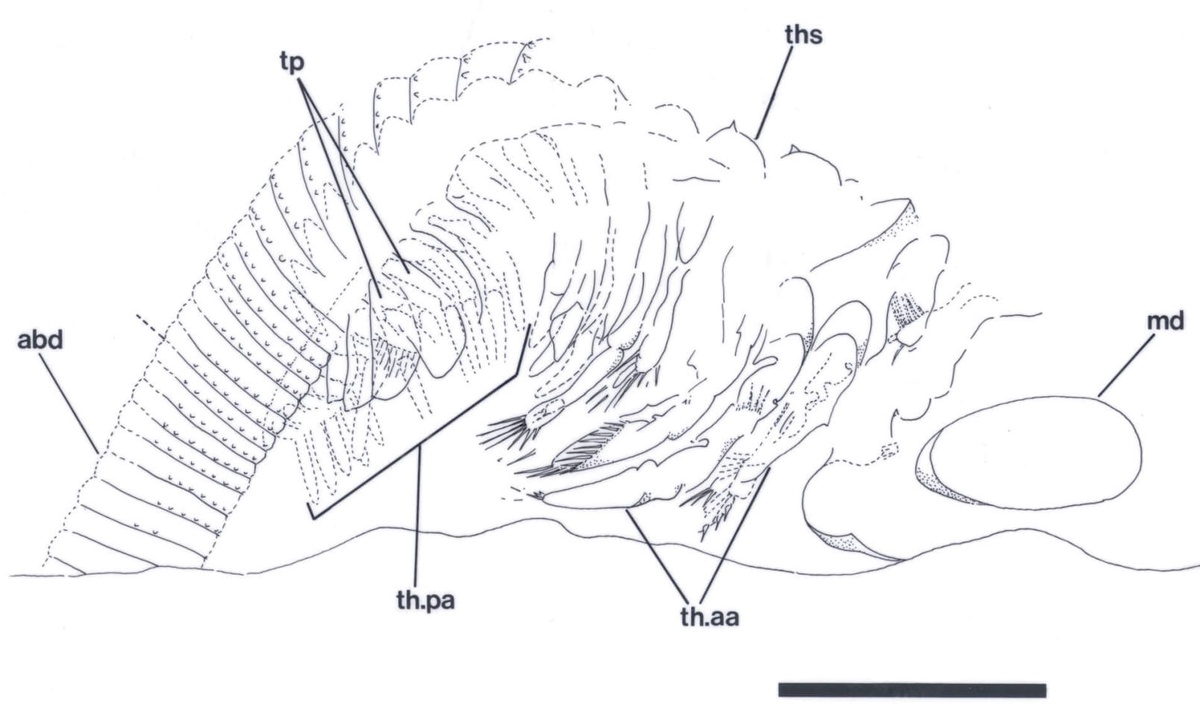
The crustacean has a large head, possibly bearing a broad cephalothoracic shield covering the thorax, with a long multi-segmented trunk ending with a telson bearing two prong-like furcal rami. The multi-segmented trunk comprises a thorax and abdomen of near identical ring-like somites (see insets below).
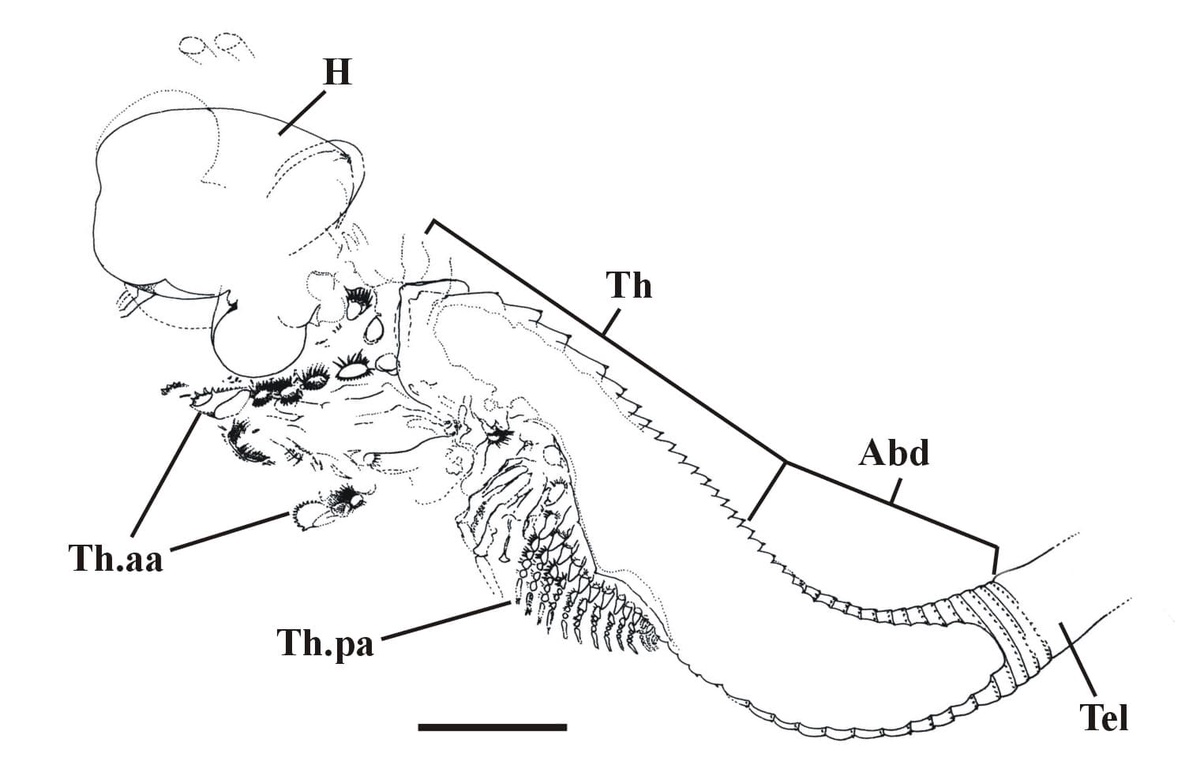
Head region
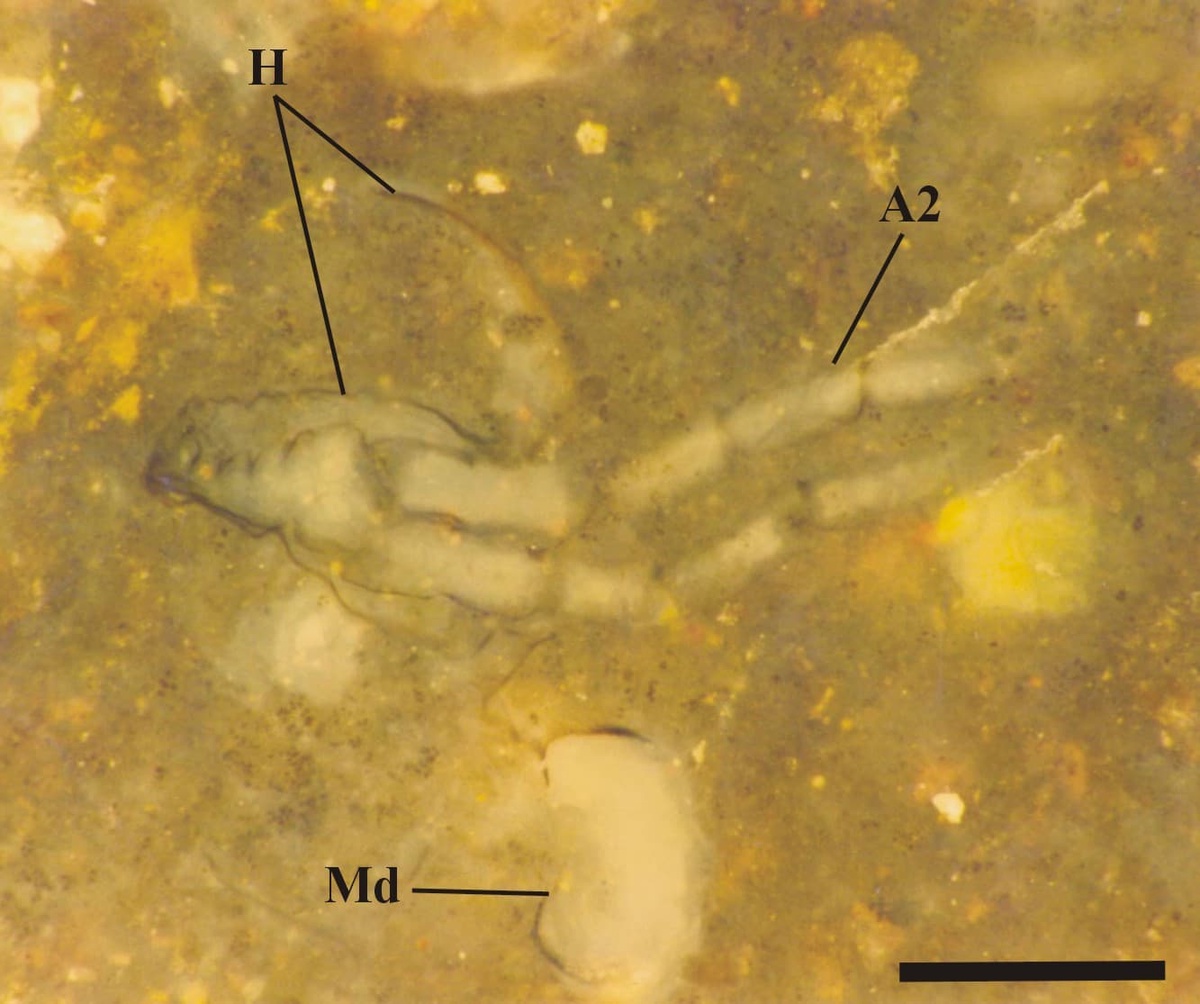
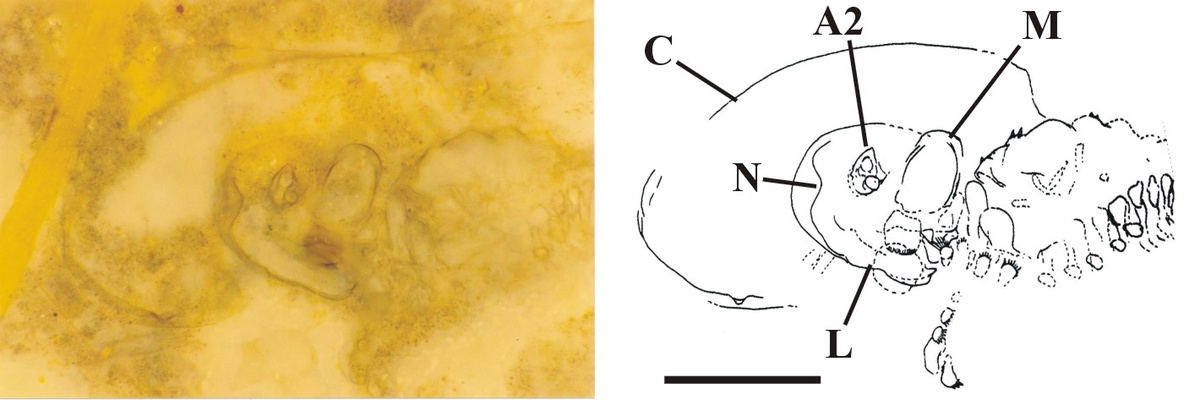
The head appears to be rounded towards the anterior where it bears a conspicuous notch, possibly representing the position of the naupliar eye (see inset below).
First antennae (antennules) have not been observed, but the second antennae are distinctive in that they are long and biramous (see inset right), with symmetrical multi-segmented rami - a feature seen in extant Onychura rather than Calmanostraca (Fayers and Trewin 2003).
Ventrally a large spatulate labrum extends posteriorly from between the second antennae and covers the mouth parts (inset below). Above this and behind the second antennae are a pair of large, robust mandibles each comprising a single coxa. The distal, mesial or grinding surface where they meet bears numerous, fine, parallel ridges. Small maxillulae are present behind the mandibles.
Body region
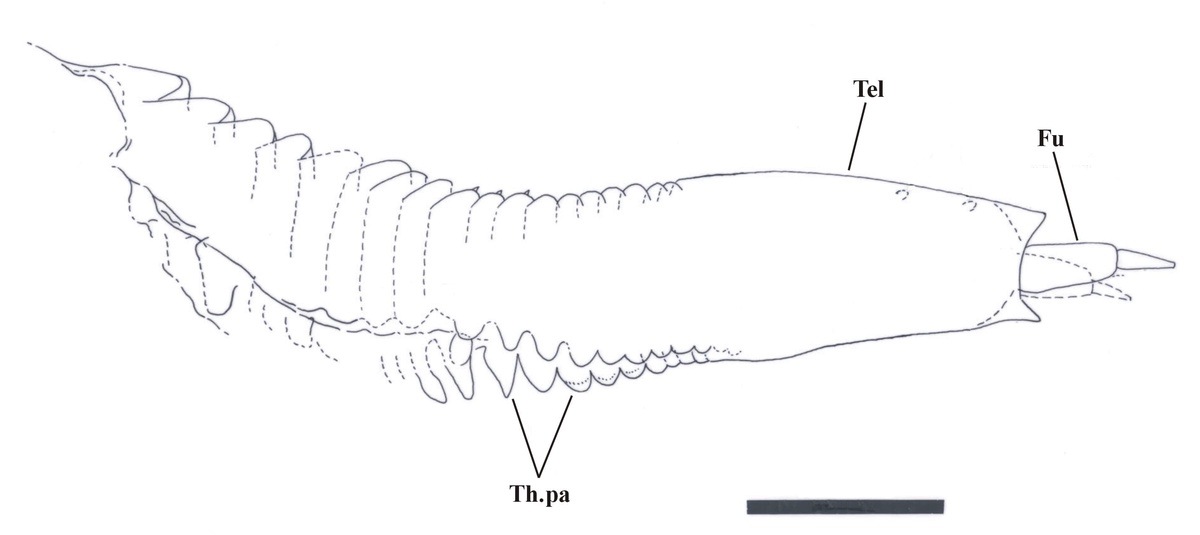
The younger growth stages of Castracollis show fewer segments on the abdomen (see inset below), and were probably added during each successive moult.
Tail region
The tail of Castracollis comprises a telson with a pair of furcal rami (see insets above and above right). The telson is oblate to cylindrical and bears two, longitudinal, sub-parallel rows of spurs on the dorsal surface. The furcal rami are attached to the posterior of the telson where they are surrounded by a series of small spurs. Each ramus comprises a single, long spinose article terminating in an articulated spine.
Leg appendages
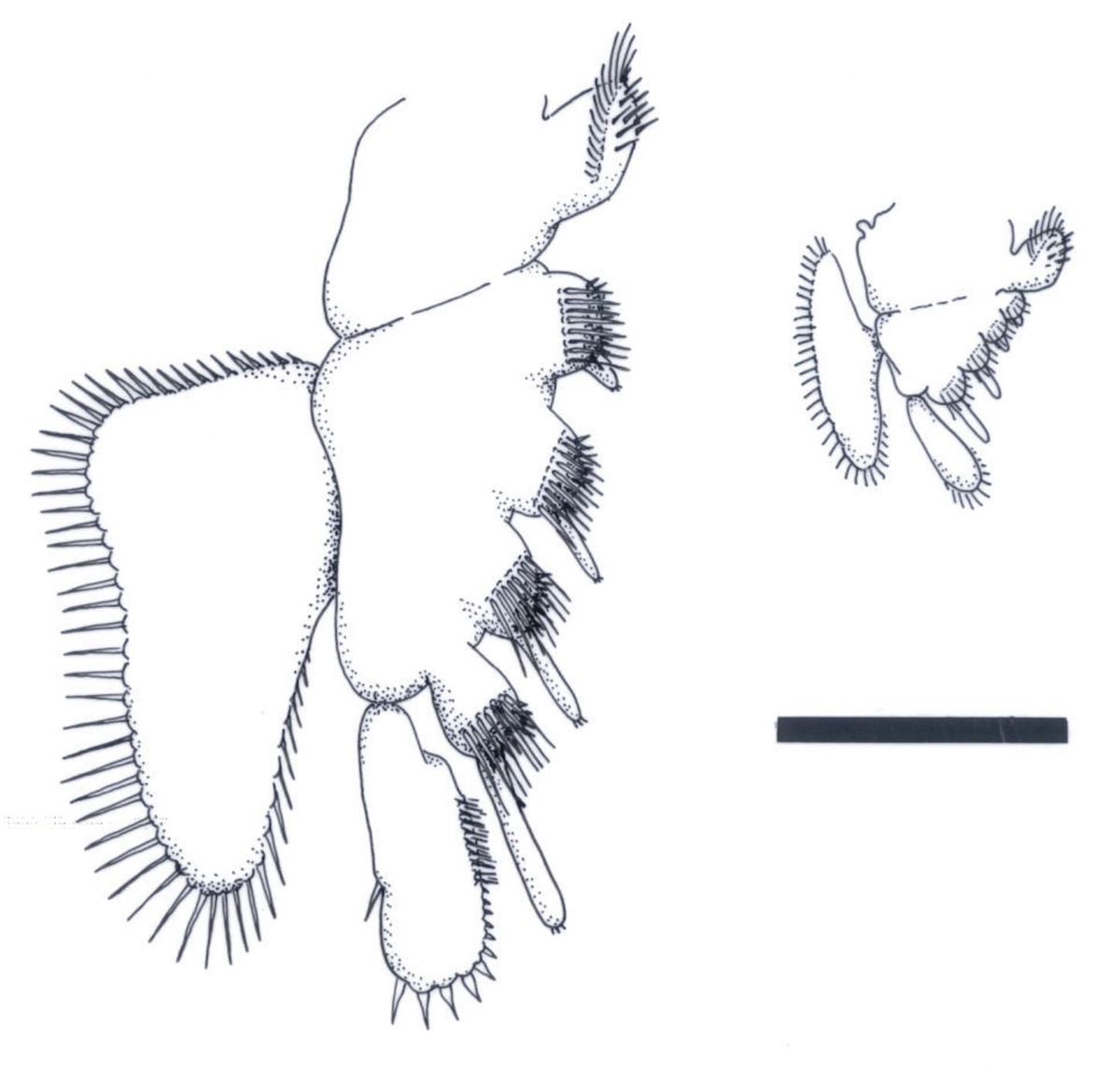
The anterior series consists of 11 leg pairs, up to 1.5mm in length and each of similar morphology (see insets right and below). Each leg comprises a basipod, bearing on the inner (mesial) surface 5 lobate endites with an array of comb-like setae. The distal 4 endites also bear a single, long tubular spine. Distally the basipod bears an articulated, raptorial and paddle-like endopod fringed with sharp spines and setae. Laterally, the distal portion of the basipod also bears a large, triangular, sheet-like exopod fringed with a row of long, fine setae.
The endopods would have been used in tearing up food items, and the setose endites used for moving the food up towards the mouth. The sheet-like exopod was probably used in locomotion. An epipod (expected to be positioned proximal to the exopod, and used as a gill) has not been observed on any of the specimens.
The posterior series of legs, of which there are variably between 10 and 15 pairs with a disposition of one pair per segment, are similar to the anterior series in their morphology, but are less than half to two thirds the length of the latter and far more delicate in their appearance (see inset right). These gradually decrease in length towards the posterior of the thorax (see insets below).

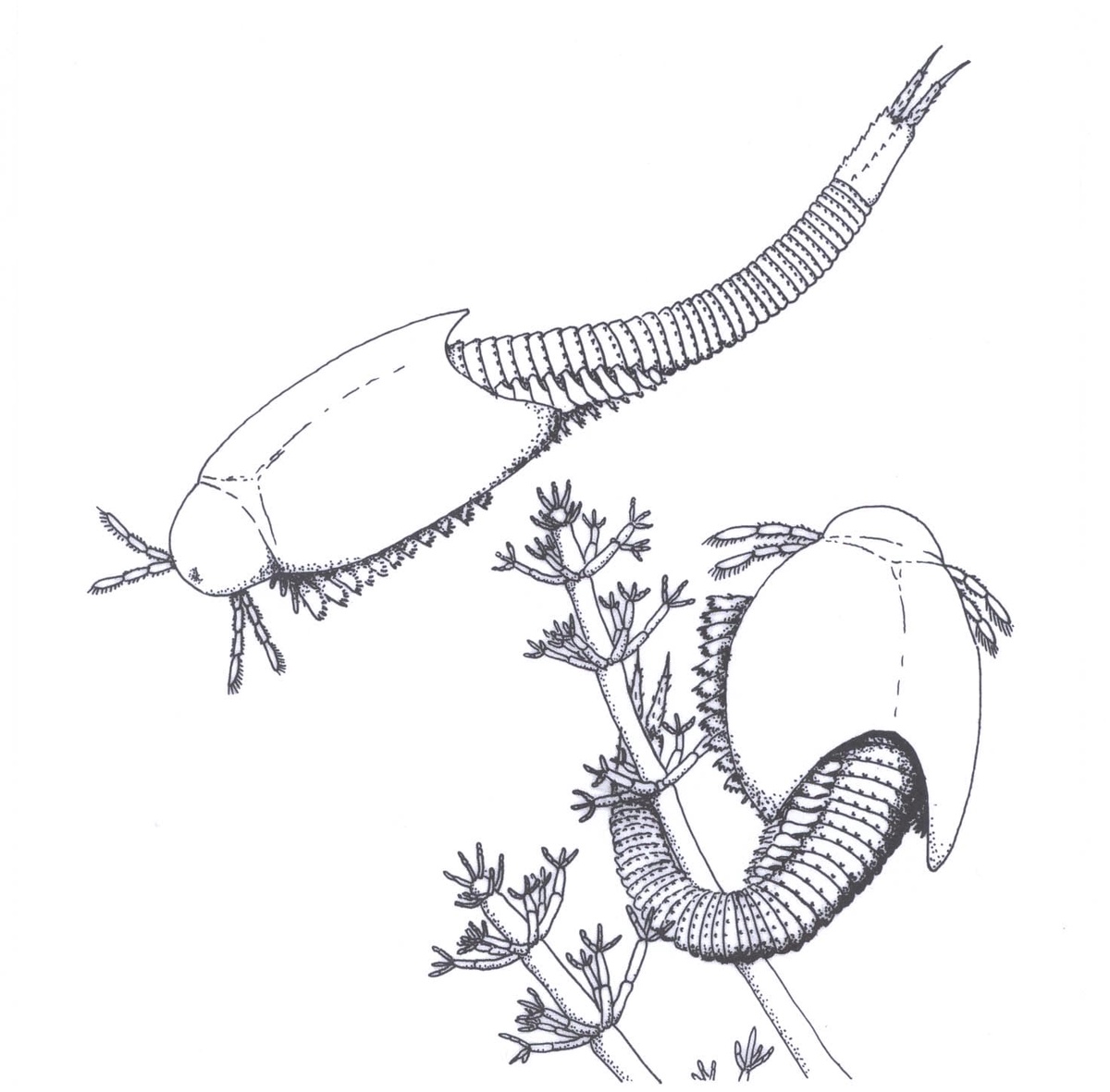
- Reconstruction
-
- Palaeoecology
-
Castracollis was undoubtedly an aquatic organism, indicated by the morphology of the leg appendages and their similarity to those of extant notostracans, namely being foliaceous with sheet-like exopods. The associated biota of Lepidocaris, various multi-cellular, filamentous and unicellular cyanobacteria together with charophytes indicate a freshwater environment.
The chert matrix that hosts the Castracollis material displays a clotted texture of aggregates of amorphous organic matter, coprolites and the filamentous meshworks. These textures are strongly reminiscent of mulm, the fluffy, amorphous organic-rich material found in many modern freshwater ponds (Anderson and Trewin 2003; Fayers and Trewin 2003).
Like modern notostracans, Castracollis appears to have been predominantly a detritivore and probably also a facultative predator. It may well have had a short life cycle, similar to notostracans, developing and moulting rapidly in temporary freshwater ponds.