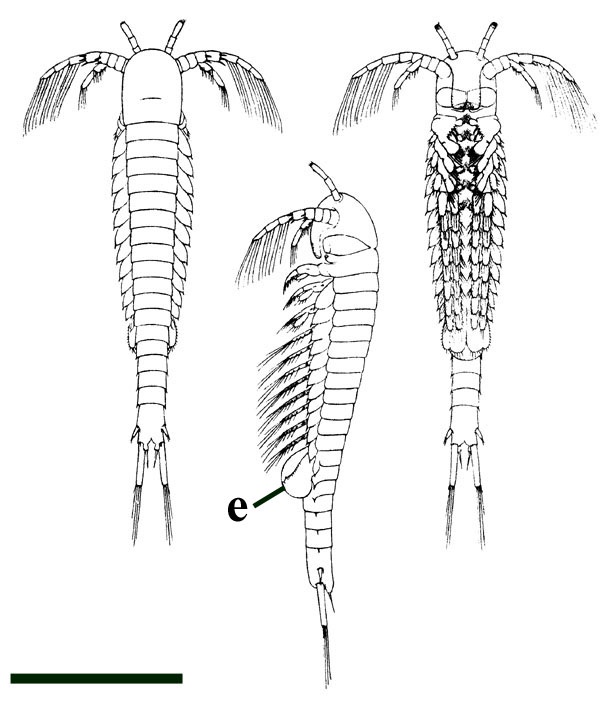
One of the better-known faunal elements from the Rhynie chert, Lepidocaris rhyniensis (see inset below) was originally described by Scourfield in 1926. Until recently, Lepidocaris or 'the beautiful shrimp', was the only crustacean known from the chert. It is the only species of an extinct order of branchiopod crustaceans called the Lipostraca and its like has not been found anywhere else than in these Early Devonian rocks at Rhynie. The relationships between Lepidocaris and other branchiopods, its morphology and palaeoecology are outlined below.
- Relationships
-
As a branchiopod, the lipostracan Lepidocaris is related to extant forms such as the tadpole shrimp Triops, the water-flea Daphnia, the clam shrimp Cyzicus and the fairy shrimp Artemia. It is an important fossil because occurring as it does in the Early Devonian Rhynie chert it represents one of the earliest known freshwater crustaceans. As such, an understanding of this early branchiopod may give us more of an insight into the timing and mechanism by which these crustaceans adapted to a continental freshwater habitat.
There are a number of differences which help us distinguish this fossil lipostracan from the other four main orders of living branchiopods. Superficially Lepidocaris is most similar to fairy shrimps, the anostracans, Scourfield (1926) noted fifteen points of difference. The following is a list some of the more important distinguishing features between all five branchiopod orders:
Lipostracans e.g.: Lepidocaris
Notostracans (tadpole shrimps)
Cladocerans (water fleas)
Conchostracans (clam shrimps)
Anostracans (fairy shrimps)
No cephalic shield in adults Single, broad cephalothoracic shield covering head and thorax Single cephalothoracic shield enclosing body but not head Bivalved cephalothoracic shield enclosing body No cephalic shield in adults (present only in hatching nauplii) Biramous 2nd antennae Reduced uniramous 2nd antennae in adults Biramous 2nd antennae Biramous 2nd antennae Uniramous 2nd antennae 11 pairs of trunk limbs, of which the first pair are modified maxillae 35 - 71 pairs of trunk limbs; 29 - 52 of which are post genital 4 - 6 pairs of trunk limbs 10 -32 pairs of trunk limbs; 1 - 16 of which are post genital 11 - 19 pairs of trunk limbs - Morphology
-
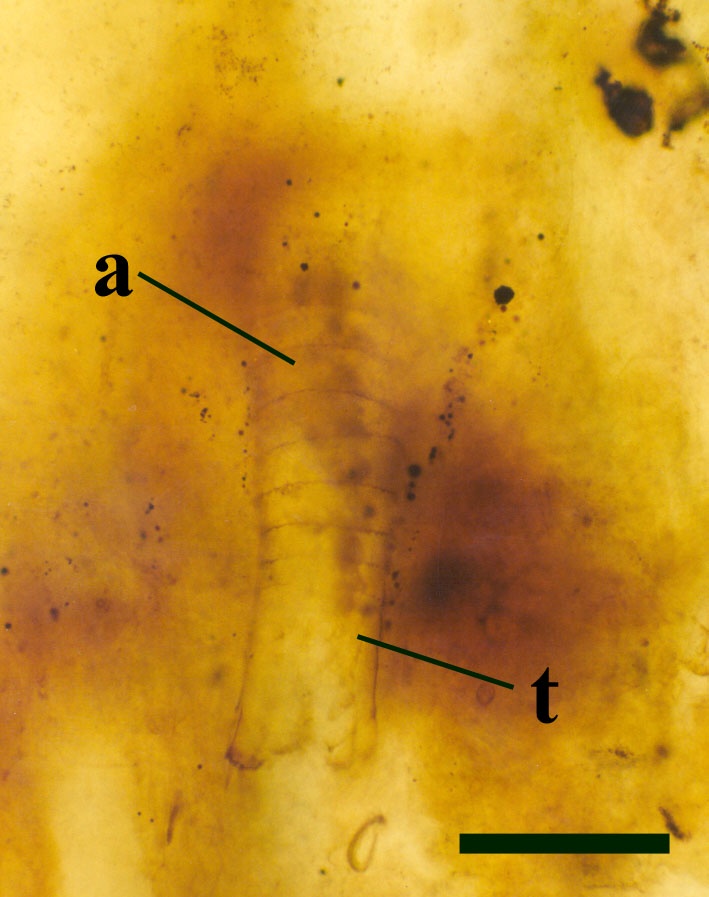
The body of Lepidocaris can be broadly divided into three parts; a distinct head region, a body or 'trunk' and a tail. No cephalic shield is present, except in the youngest stages (Scourfield 1940c). In many specimens the fossils appear disarticulated or comprise isolated fragments or clusters of material. This suggests that many of the specimens probably represent exuviae or moults.
Forms in various stages of development have been found from probable egg cases, through juvenile stages (Scourfield 1926, 1940c) to fully grown adults. Adults attained a maximum length of 3mm. Consequently we are able to establish much of the ontogeny of Lepidocaris. Both females and males have also been identified.
Head Region

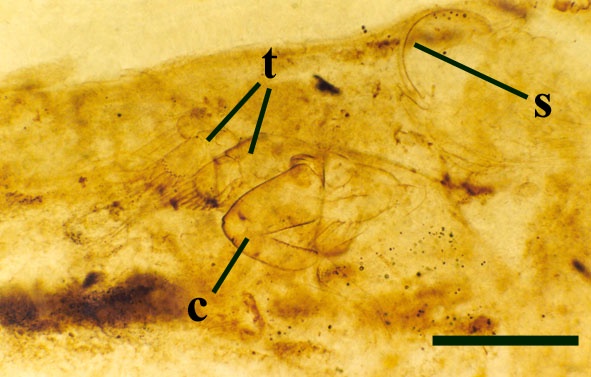
The head lacks eyes, but possesses two pairs of appendages, the first antennae (or antennules) and second antennae, anterior to the mouthparts. The first antennae are small, and uniramous comprising three segments or articles, the most distal bearing very fine setae or 'hairs'. The second antennae are characteristic, being long and biramous (see inset right).
The main 'branch' comprises eight articles, the three most distal articles bearing long plumose setae. The second 'branch' is attached to the third proximal article of the main branch and comprises two or three articles in female and male forms respectively. The mouthparts comprise a pair of robust mandibles (see inset right) and posterior to these the maxillulae. In females the maxillulae are small, consisting of a single plate bearing a single spine and rows of setae.
In males, however, these are developed into large clasping organs (see inset below and below right). The maxillae are greatly modified and 'leg-like' (Walossek 1993), forming what Scourfield (1926) originally described as the first pair of trunk limbs (see insets below).
These clearly belong to the head region, because the corresponding segment is fused with the head capsule. The junction between the maxillae segment and the rest of the head capsule is marked by a 'mandibular groove' (see reconstruction towards top of page). The osmoregulatory 'neck organ', seen in extant branchiopods, and a shared feature or synapomorphy of Branchiopoda and Maxillopoda (Walossek 1995), has not been observed in Lepidocaris.
Body Region
The body or trunk of Lepidocaris comprises a thorax and an apodous abdomen. Excluding the head and tail, the trunk in adult forms comprises seventeen segments or somites. The first fourteen ring-like somites, following the back of the head capsule, bear lateral scales. The final three somites form apodous, simple rings. Lepidocaris has ten pairs of foliaceous trunk limbs with a disposition of one pair per somite. These form a complex filter feeding system, a feature which, together with the presence of a sternal food-groove, is characteristic of the Branchiopoda (Walossek 1993, 1995) The first two pairs of trunk limbs are similar in morphology to the maxillae. These and the maxillae are modified with the terminal articles (endopods) being slightly robust, palmate and comb-like (see inset below and below right).
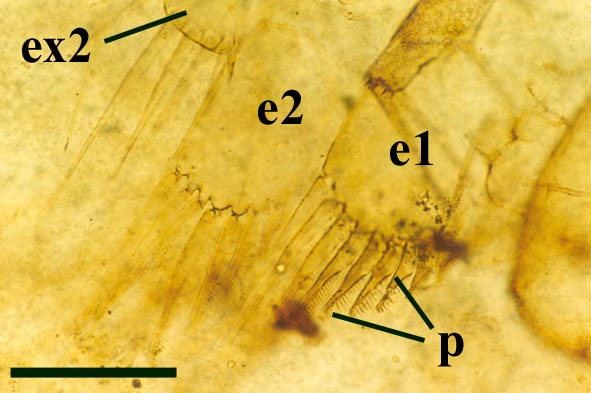
The following eight pairs of appendages are far more foliaceous and delicate (see inset below), and are rather copepod-like in their appearance. Posterior to this series of appendages in females occurs an egg pouch and cover that may have developed from modified trunk limbs. In males, however, the last two pairs of appendages are slightly modified, the exopods and endopods appearing broader than those on the preceding appendages.
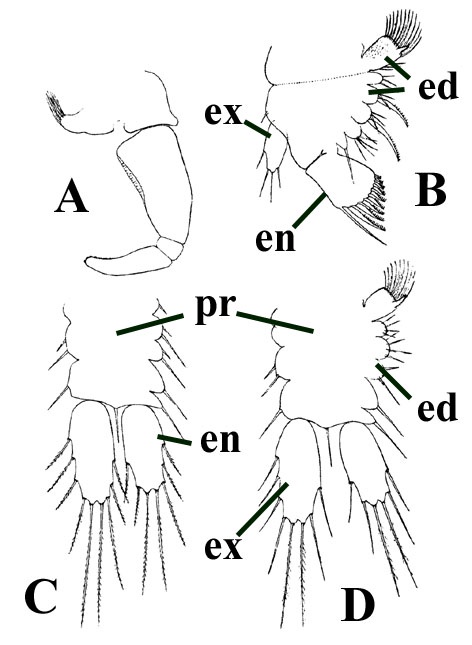
Tail
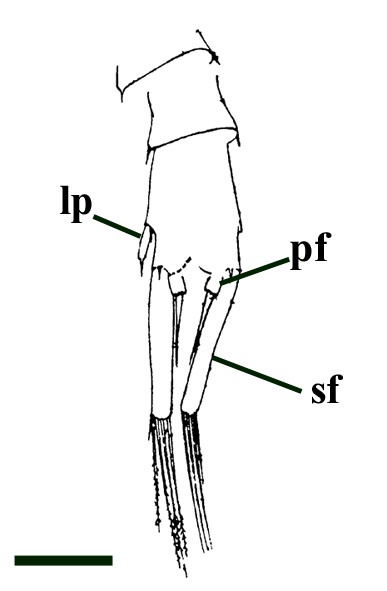
- Palaeoecology
-
Lepidocaris is perhaps one of the more abundant faunal elements found in the Rhynie chert. It is typically found associated with charophytes, coprolites, algal filaments and often occurs in cherts displaying a clotted or 'mulm-like' texture and was undoubtedly an aquatic organism. The delicate, foliaceous nature of the trunk appendages indicates Lepidocaris was an active swimmer, the more pectinate anterior trunk appendages and maxillae quite probably being used for feeding and food gathering.
The general ecology of Lepidocaris may have been somewhat similar to that of modern anostracans, perhaps thriving in small temporary ponds, feeding off algae and organic particulates. The common association of the animal with the charophyte Palaeonitella, assuming the ecology of the latter was similar to modern day Nitelleae, could suggest Lepidocaris, as with many modern anostracans, preferred to inhabit alkaline waters.
One final point can be made about the palaeoecology of this enigmatic branchiopod in comparison with modern anostracans. Based mainly on the mixture of primitive and modified features Scourfield (1926) observed in Lepidocaris, together with the overall lack of contemporaneous aquatic invertebrates in the chert, he suggested that the animal must have adapted to live in a 'very peculiar' environment. In fact he postulated the lipostracan may have adapted to live in hot, silica-charged water. However, there is no evidence to suggest that prior to silicification temperatures in these inhabited pools were any different to similar freshwater environments elsewhere, particularly considering the aquatic flora present.