Mites are from the second largest order of the arachnids called the Arcari, an order that also includes the ticks. These arthropods are generally very small in size very often being less than a millimeter in length, but the largest trombiform mites can reach up to 12mm in size. Mites range from the Early Devonian to the present day. The earliest member of this group comes from the Rhynie chert and was originally described by Hirst (1923). The fossil record of mites is particularly sparse and like most other arachnids they occur mainly in sites of exceptional preservation where their soft unmineralised cuticle can become fossilised. Although others are known from the Devonian (e.g. Norton et al. 1988) the majority of fossil mites are best known from the Oligocene Baltic amber.
- Relationships
-
As with other arachnids (including true spiders, harvestmen (opilionids), trigonotarbids and scorpions), mites are part of a group of animals called the chelicerates, which also includes horseshoe crabs (e.g. the modern Limulus) and the extinct eurypterids. Although superficially similar to spiders and harvestmen there are however a number of characteristic differences that separate them; the two main differences are tabulated below:
Mites Spiders Opilionids (harvestmen) Lack distinct division between head and body. Clear division into a head and body. Lack distinct division between head and body. Smooth opisthosoma. Smooth, fused opisthosoma (with the exception of liphistid spiders). Segmented opisthosoma. - Identity
-
One species of mite was originally recorded from the Rhynie chert, Protacarus crani (Hirst 1923). Since then Dubinin (1962) transferred four of the specimens originally assigned to Protacarus to four new species: Protospeleorchestes pseudoprotacarus, Pseudoprotacarus scoticus, Palaeotydeus devonicus and Paraprotocarus hirsti. For the purposes of this resource page, however, we will concentrate on the first described species, Protocarus crani.
Protacarus has been placed as a member of a sub group of the Arcari, the family Pachygnathidae which still survives today. This is a group of primitive soft-bodied mites of which the extant forms commonly live in soil and damp forest litter.
Note: When the specimens of Protacarus crani and the other Rhynie mites were first prepared they were found by fragmenting small blocks of chert into tiny shards. The shards of chert that were found to contain the mite were then mounted onto glass slides. Since these fossils are in uneven shards of chert and often preserved in 3D, they cannot be easily photographed with any clarity at high magnification, therefore most of the images seen on this page are taken from line drawings after Hirst (1923) and Dubinin (1962) (e.g.: see inset under Morphology below right).
- Morphology
-
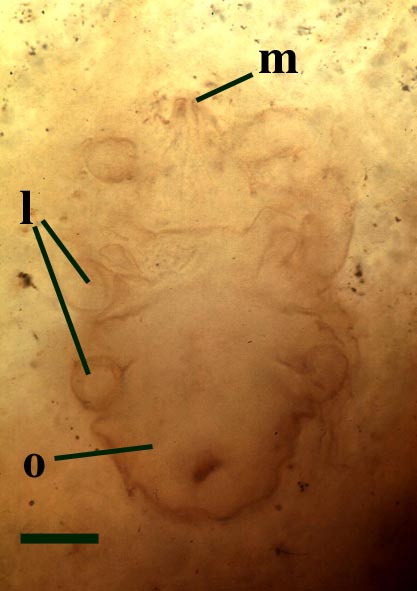
The Rhynie mite Protacarus has a characteristic and distinctive morphology that easily distinguishes it from other Rhynie arachnids, namely the trigonotarbids. Overall, mites including Protacarus show no segmentation on the opisthosoma and no distinct division between the head and the rest of the body. The size is minute, the fossil mites found ranging from 290 to 450 microns in length (see insets below).
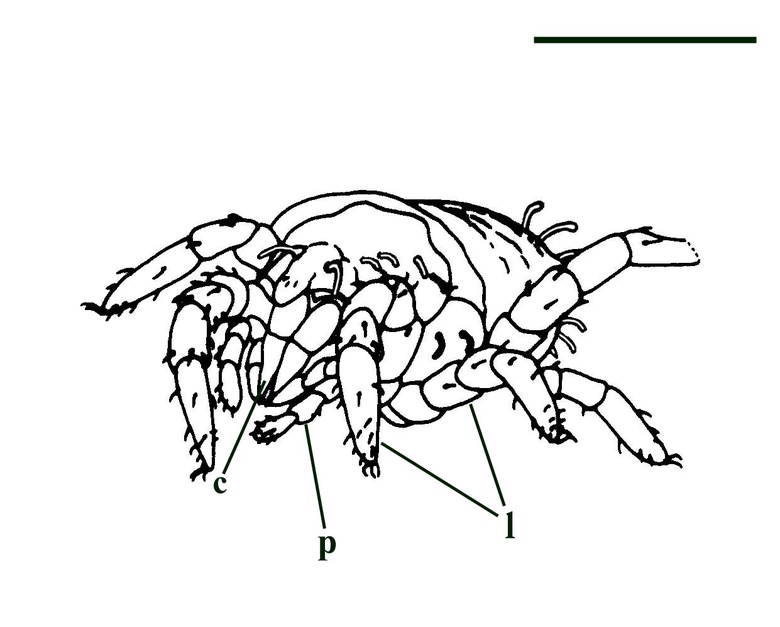
Protacarus does not exhibit the same scale-like cuticle sculpture as that seen in the Rhynie trigonotarbids. The body is divided into an anterior proterosoma and a posterior hysterosoma, the junction being taken in front of the posterior two pairs of legs. The proterosoma is subdivided into the gnathosoma (bearing the mouthparts, chelicerae and pedipalps) and propodosoma which bears the first two pairs of walking legs. Protacarus lacked eyes. The hysterosoma is broadly subdivided, comprising the metapodosoma (the region bearing the last two pairs of walking legs) and the opisthosoma. The hysterosoma exhibits transverse furrows and the dorsal surface exhibits very fine fan-like hairs or setae (see inset above left).
The paired chelicerae ("jaws") are relatively large and highly modified, the terminal segment being strongly cutinised, having a pointed tip and appearing very stylet-like. Lateral and posterior to the chelicerae are the pedipalps. These comprise six jointed segments or podomeres with three claw-like hairs on the terminal segment (the tarsus). There are four pairs of walking legs, each comprising just six jointed segments. Again the walking legs possess claw-like hairs at the tips of the terminal segments. The walking legs and the pedipalps show very fine hairs or setae which most likely formed part of the animal's sensory array (see inset above left).
- Palaeoecology
-
The extant Arcari are particularly diverse in terms of their structure and habitat. They are the only arachnid order which contains plant-eating species.
A number of species are parasitic, and several are aquatic; occurring in fresh-water and even marine habitats. The mites that include the family Pachygnathidae are in part saprophagous (living off dead organic remains) and mainly found living in damp plant litter; it is quite possible that Protacarus lived in a similar habitat.
Many of the Rhynie mites have been found within empty sporangia. The morphology of the mouthparts of Protacarus suggests it may have also fed off the sap of living plants by puncturing the epidermis with its stylet-like chelicerae then imbibing the fluid through the wound.
In some thin sections of chert, axes of plants have shown pathological damage and what appear to be enlarged cells in the epidermis and cortex filled with opaque organic material that may reflect a response to such an attack (Kevan et al. 1975).
- Other useful references
-
For a review of the phylogenetic importance of the Rhynie chert mites see Bernini (1986).