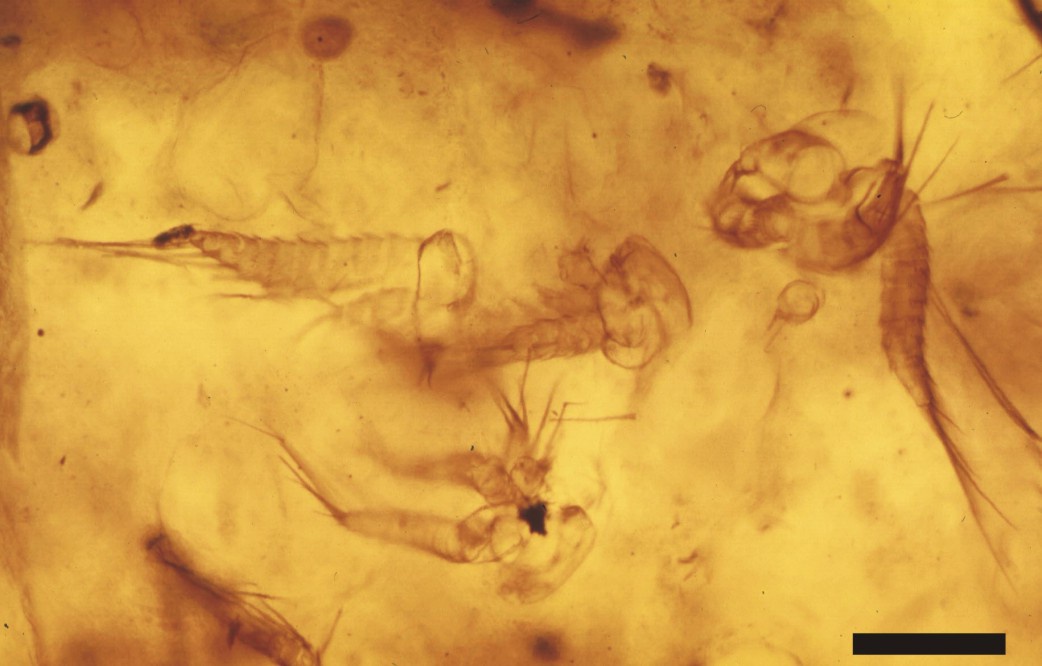
As well as the Lepidocaris rhyniensis, Scourfield 1926, the Windyfield chert has sourced other crustacean remains, namely numerous specimens of branchiopod nauplii. All have been found within clotted and coprolitic chert textures, almost exclusively within brecciated, nodular cherts. These are currently being described for publication in the scientific literature.
- Occurrence
-
The branchiopod nauplii have been found either as aggregates of numerous individuals (see inset at top of page) or as isolated specimens (see inset above), in close association with very well preserved specimens of the charophyte Palaeonitella cranii, and suspended within clots and cumuli of amorphous organic matter and coprolites.
The organic-rich clots and cumuli, faunal elements and charophyte remains locally form discrete aggregates, which are in turn enveloped by meniscate mucilages of microbial filaments crossed by fungal hyphae. The association of charophytes, coprolites, and a clotted organic-rich matrix with these crustaceans, together with their morphology (see below) indicates these were freshwater organisms.
The clotted and filamentous organic framework appears open, and the remarkable preservation of the charophytes, fungi, coprolites, and the crustaceans indicates that silicification occurred whilst the material was still in an aquatic environment.
- Morphology
-
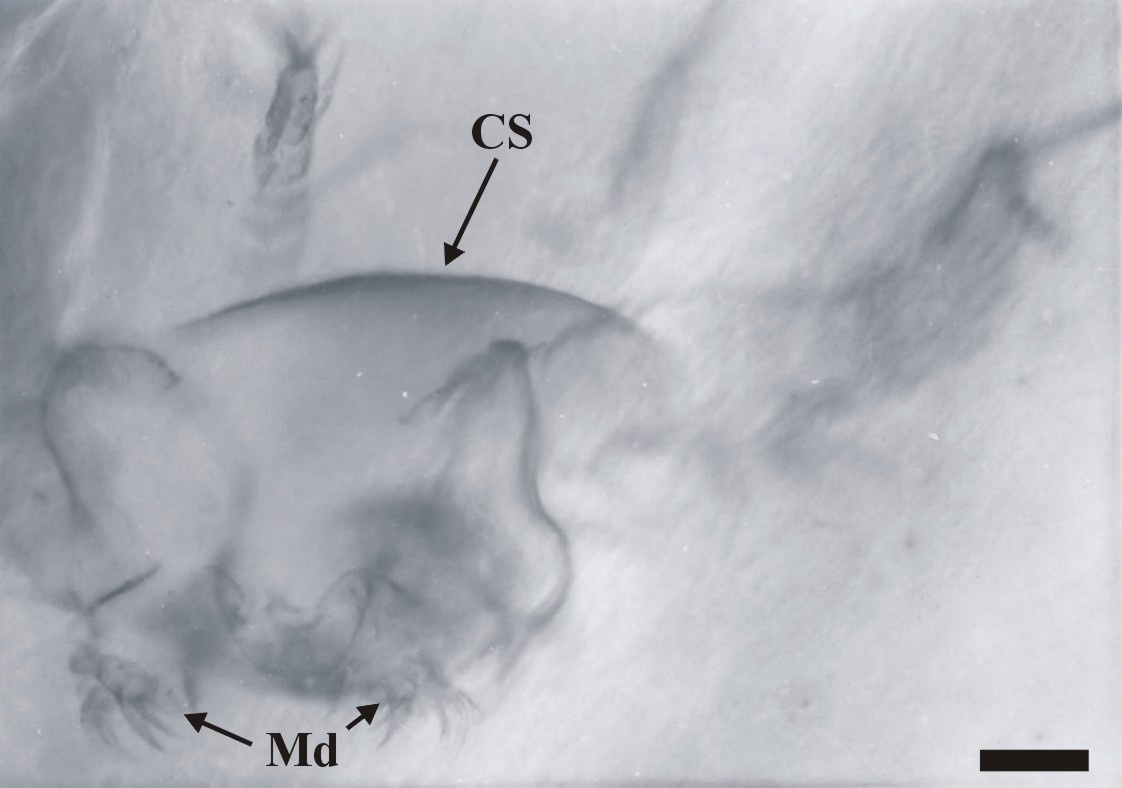
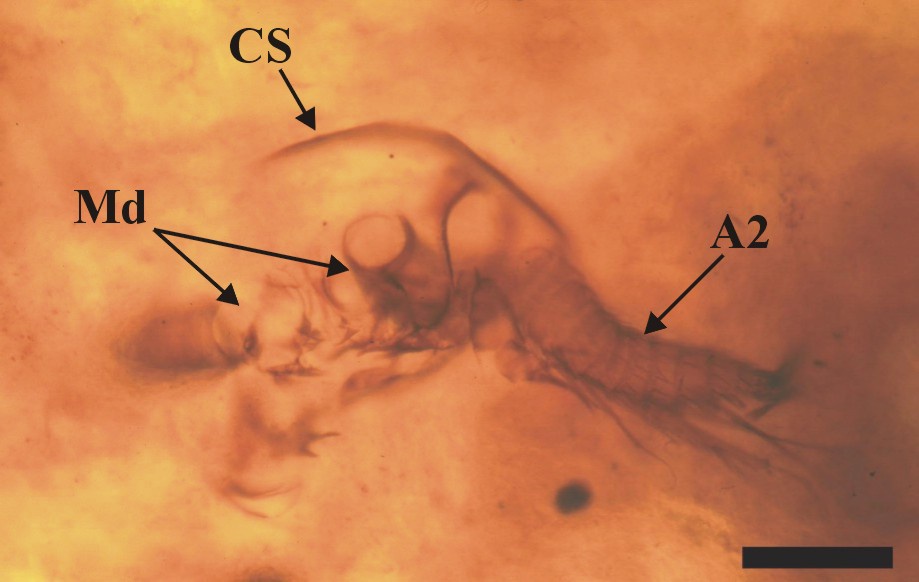
The total length of the nauplii has not been fully ascertained, mainly due to the orientation of the specimens in the thin sections, and also because of their otherwise fragmentary nature, but on the whole it appears to be at least 300mm. Excluding the length of the appendages, the body width is up to 250mm, corresponding to the width of the cephalic shield (CS).
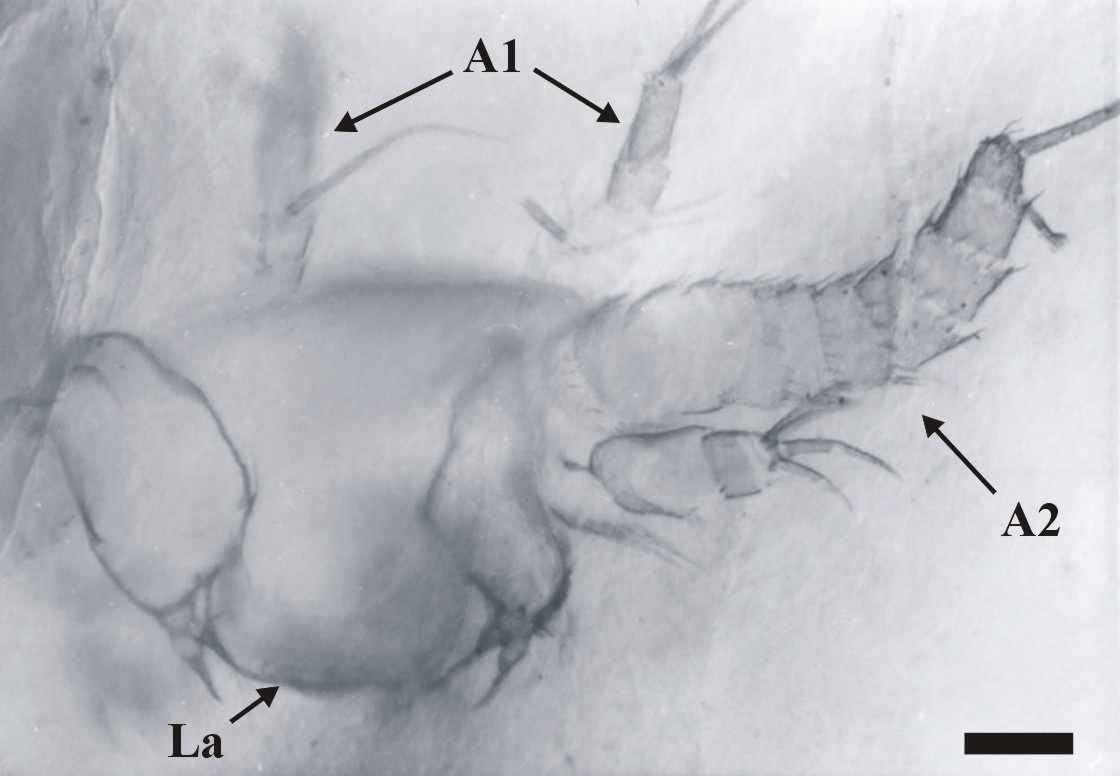
The dorsal area of the cephalic region comprises a broad, dome-like cephalic shield, made of very thin, smooth cuticle (see insets above). The anterior of the head has not been fully discerned, but it generally appears to be rounded. Ventrally a broad labrum (La) extends from between the position of the second antennae (A2), posteriorly towards, and overlapping the position of the mandibles (Md).
Appendages
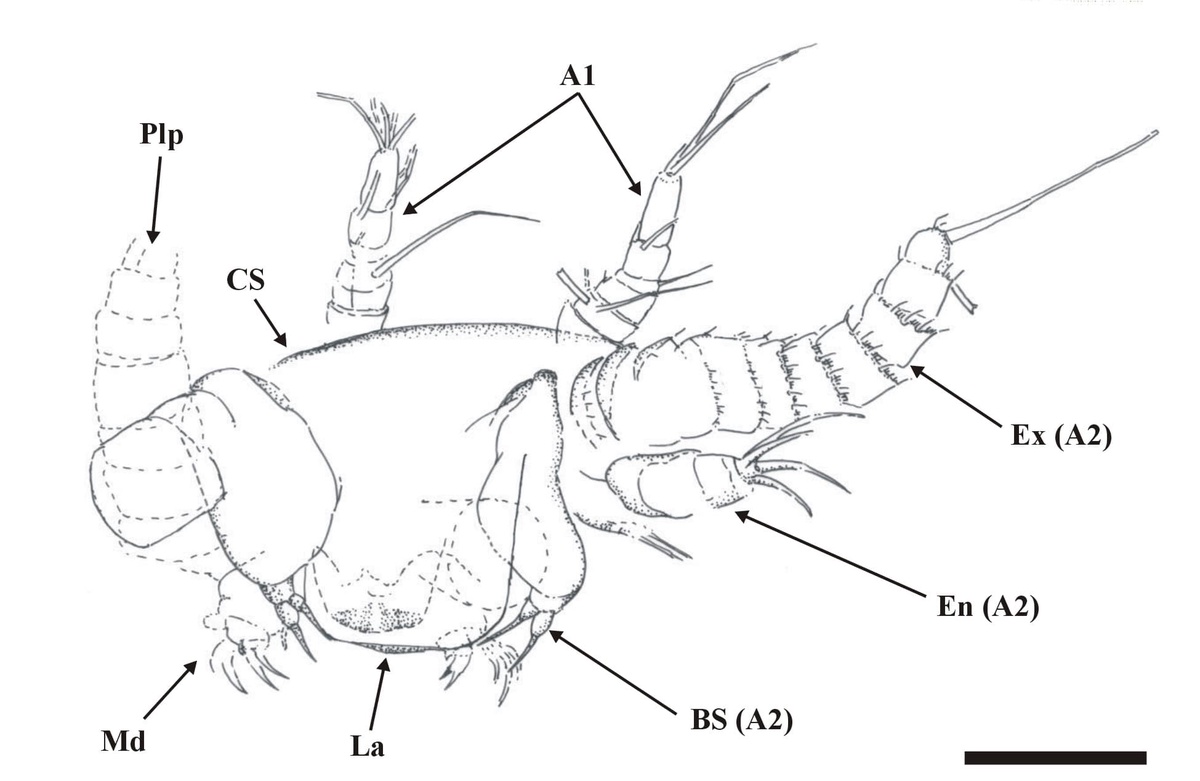
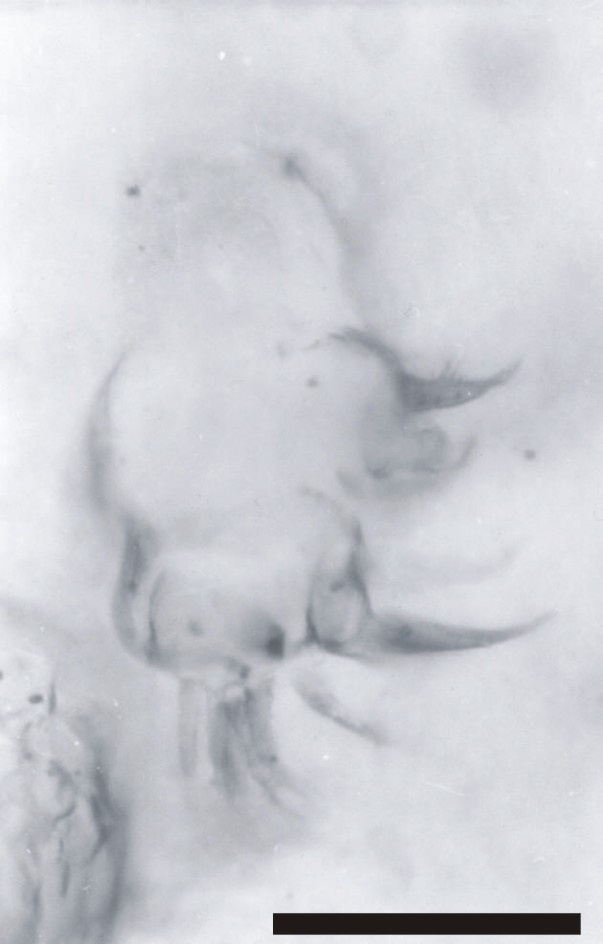
Immediately posterior to the first antennae are the second antennae (A2), each comprising a long multi-segmented outer ramus or exopod (Ex), a smaller endopod (En) and a large proximal endite with bifid seta (BS) (see inset above). The multi-segmented exopod comprises between eleven and twelve tubular segments of similar morphology. The most distal article is rounded or blunted and bears up to three long terminal setae, up to 250mm in length. All the other articles possess a single, medially positioned, long seta up to 250mm in length, which together with the three setae on the most distal element gives the exopod a very plumose appearance. The base of each long seta and the distal margin of each article display an array of smaller setae (see inset right).
The endopod (En) comprises two or possibly three articles, and is up to 100mm in length. Distally, on what may possibly be a third article, are up to four setae.
The exopod ramus appears to be articulated directly with a large proximal endite. This appears large and robust, and bears a pronounced bifid seta (BS) (see insets above).
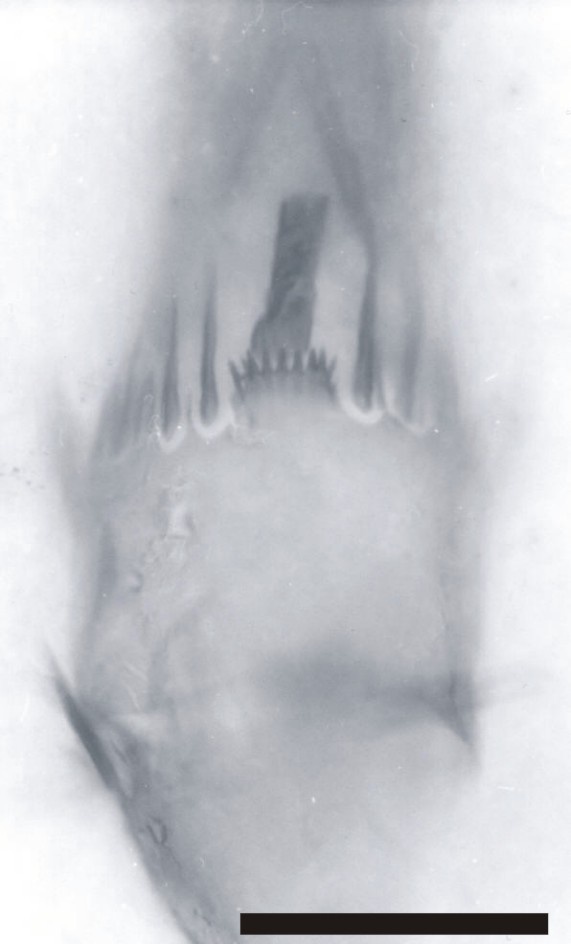
Posterior to the second antennae are the mandibles (Md), comprising a proximal endite or coxa, and an articulated palp (Plp). The morphology of the mandibular palp is similar to that of the exopod of the second antennae, forming a multi-segmented ramus, comprising at least seven to eight tubular segments. The setation of the segments is the same as that for the exopods of the second antennae.
The mandibular coxa, or proximal endite is around half the size of the proximal endite of the second antenna, and is broadly rectangular in shape. The proximal endite typically appears lobate, supporting an array of spine-like setae (see inset right).
- Identity
-
The nauplii are undoubtedly branchiopodan in their design, especially when comparing the appendage morphology to those of the nauplii of the Euanostraca, or Brine shrimp (e.g. Walossek 1993, Fig. 44A ).
The remains in the Windyfield chert are undoubtedly true naupliar or larval stages since only the first and second antennae and mandibular appendages occur, and at most only a small, undivided hind body is present. This is comparable, for example, with the first or possibly second instar of the euanostracan Artemia salina ( Walossek 1993 and references therein). Scourfield (1940c) described two young individuals of Lepidocaris rhyniensis, both showing a post-cephalic trunk, delineated into segments and bearing at least rudimentary trunk appendages.
The younger of these two individuals ( Scourfield 1940c , Fig. 2) shows six trunk appendages (not including the modified, trunk limb-like second maxilla), which equates to the tenth instar of Artemia salina. Therefore these new crustacean remains represent the earliest ontogenetic stages or instars of a branchiopod found to date in either the Windyfield or Rhynie cherts.
No post-naupliar instars or adult forms that belong to the same animal are present in the same chert samples. This suggests the nauplii may have died very early on in their development, possibly due to a sharp change in water conditions, such as the introduction of erupted hot-spring fluids into their aquatic environment.
If the nauplii are of Lepidocaris, the smaller number of articles in the exopods of the second antennae and mandibles in the post-naupliar stages could be explained by a reduction in the number of articles from the larval stage during the following ontogenetic stages.
At present, we cannot say with any conviction if the Windyfield chert nauplii are of Lepidocaris, or if they may pertain to another form of anostracan branchiopod hitherto unknown in the cherts, but the similarities observed between these nauplii, extant euanostracan nauplii (e.g. Artemia), and the known post-naupliar stages of Lepidocaris suggests they almost certainly belong in the anostracan lineage.