Collembolans or 'springtails' are a group of small arthropods that, together with the Protura, comprise the Class Parainsecta.
As with true insects the Parainsecta are hexapods in that they possess three pairs of walking legs. However they differ in the fact that the abdomen of true insects comprises eleven segments whereas the collembolans have only six abdominal segments.
The Protura on the other hand possess between nine and twelve segments on the abdomen. In the past the collembolans, proturans and two other groups; the diplurans and the thysanurans were all grouped together as the apterygotes or the wingless insects. It is now generally accepted by entomologists, however, that the first three groups are in fact offshoots of the main insectan evolutionary stem and have therefore since been given separate taxonomic status from the Class Insecta.
Thus the frequently repeated claim that the earliest known 'insect' is found in the Rhynie chert refers to a collembolan, and the 'insect' taxonomic status depends on taxonomic opinions.
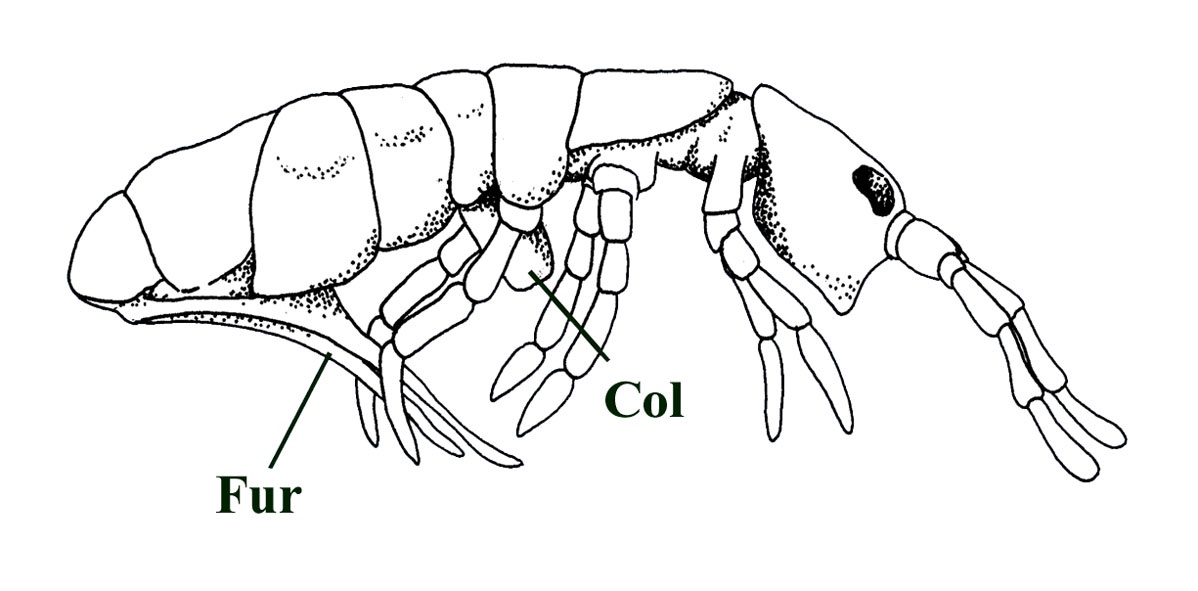
The collembolans (with the exception of a few genera) are also distinguished by a curious ventral abdominal 'jumping organ' - the furcula which the animal uses to vault into the air when it is threatened - hence the common name 'springtail'.
This is attached to the fourth abdominal segment and is held in tension by means of a special 'catch' called the tenaculum on the third abdominal segment. Another curious characteristic of collembolans is the collophore, a ventral tube attached to the first abdominal segment.
The exact function of this organ remains unknown but may be a means of allowing the animal to regulate it's water balance. The collophore was originally thought to be used for adhesion - hence the name 'collembolan' taken from the Greek words coll and embol meaning 'glue peg'.
- Fossil Record
-
The fossil record of the collembolans and indeed the proturans is particularly sparse, in fact proturans have not been described from the fossil record at all. The collembolans have been in existence since the Early Devonian; with most fossil representatives having been described from Oligocene amber but the earliest recorded fossil species is from the Rhynie chert.
To date one species has been identified; Rhyniella praecursor, described by Hirst and Maulik (1926) (from four specimens of the animal's head) and is traditionally famed for being the earliest recorded fossil 'insect'. Initially its affinity was treated with some skepticism, however Tillyard (1928), demonstrated that Rhyniella was indeed a collembolan based on the morphology of the antennae and mouthparts.
Further discoveries of more complete specimens by the Rev. W. Cran and D. J. Scourfield established beyond a doubt a collembolan affinity (Scourfield 1940a, b). Rhyniella shows very similar characteristics to two extant collembolan families; the Hypogasturidae and the Entomobryiidae. The similarity is so striking, it has lead some authors in the past to question whether the Rhyniella specimens are modern contaminants (Crowson 1970), but since they are all completely silicified and enclosed within the rock, they are undoubtedly contemporaneous with the chert and are therefore Devonian in age.
Although strictly speaking the collembolans are no longer regarded as true insects by most entomologists, this in no way detracts from the significance of this fossil in terms of the evolutionary history of hexapods and the insects and also the emergence and construction of early terrestrial biotas.
Note: When the specimens of Rhyniella praecursor were first prepared in the 1920's they were found by fragmenting small blocks of chert into tiny shards. The shards of chert that were found to contain Rhyniella were then mounted onto slides. Since these fossils are in uneven shards of chert, they cannot be easily photographed with any clarity, therefore the following images on this page are taken from line drawings after Hirst and Maulik (1926) and Scourfield (1940a, b).
- Morphology
-
The general morphology of Rhyniella praecursor appears very much similar to the general body plan of many modern collembolans. It was rather small, apparently being between 1-2mm in length.
The head, thorax (the region that bears the walking legs) and parts of the abdomen have been identified and described (see inset below). The body segments behind the head do not appear to have been fused together and were sub-cylindrical. Work on new specimens by Whalley and Jarzembowski (1981) showed that Rhyniella possessed a furcula. The following gives a brief account of the morphology of the known body elements.
Head Region
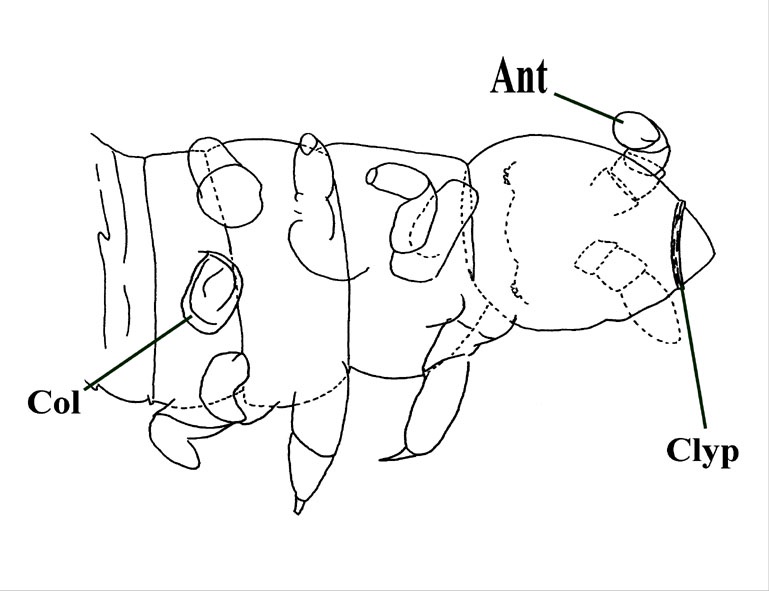
The head varies between 300 and 400 microns in length and is heart-shaped with the pointed end to the anterior. It has an arched strongly cutinised clypeus commonly appearing in specimens as a conspicuous dark brown band (see inset below left).
A labrum is present (a small ventral piece of cuticle that protects the mouthparts) attached to the clypeus by two small rounded projections and appears to have been covered by tufts of fine hairs or setae (see inset below right). A pair of ocelli or 'eyes' are present, each apparently comprising at least six tiny hemispherical tubercles on the dorsal part of the head behind the antennae.
The antennae are simple, rather shorter in length than the head capsule, each comprising four segments. The terminal segment may possess tiny sensory structures.
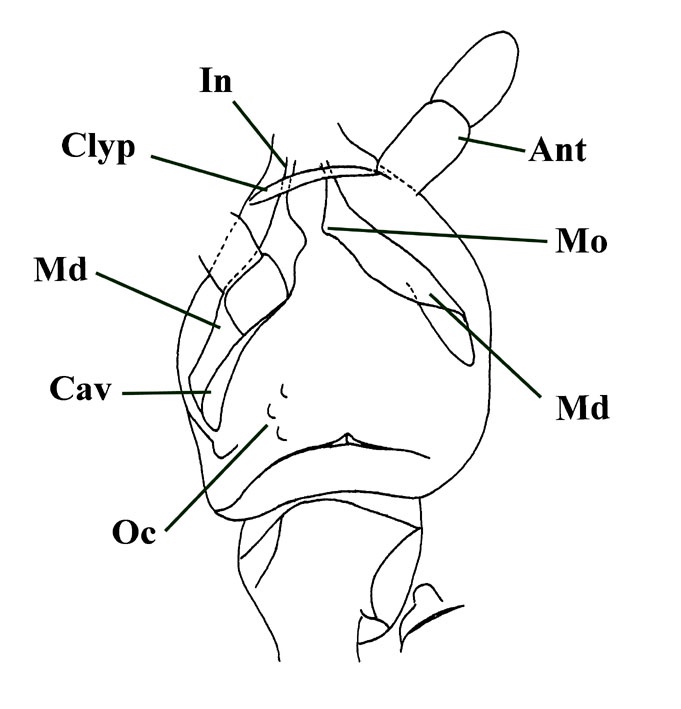
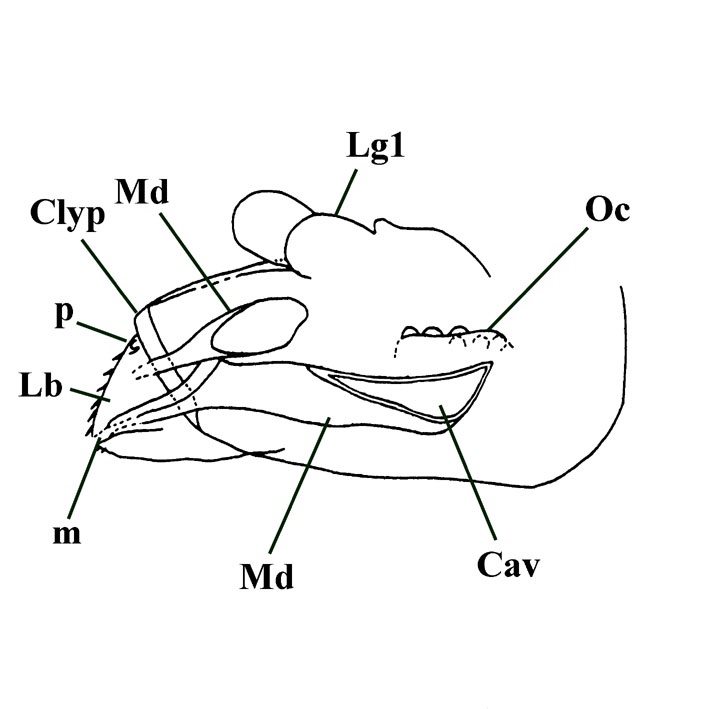
The mouth parts are of interest. These are entirely located within the head capsule. The most conspicuous feature are the mandibles. These are relatively elongate (see insets above left and right) and become split towards the anterior into a distinct incisor and molar region.
The tips of the incisors have not been seen but were probably stylet-like and most likely lacked teeth. In one particular specimen the molar area of one mandible shows very faint parallel lines. This suggests the molar area originally supported some very fine ridges as 'teeth'.
The posterior end of the mandibles show a conspicuous cavity into which muscles were originally inserted. Other mouthparts are unfortunately not particularly clear, though a number of specimens show what may be parts of the maxillae and possible lingual elements.
Thorax and appendages
The thorax is the region of the body between the head and the abdomen that supports the walking legs, and may be divided into three parts; the prothorax, mesothorax and metathorax. The prothorax bears the first pair of walking legs.
In Rhyniella this segment is not very clear and seems to be very much reduced and lacking a tergum giving the impression that the first pair of legs originates between the back of the head and the following segment (the mesothorax). The mesothorax and metathorax are well developed (see inset below); these two segments bearing the second and third pairs of walking legs respectively.

The legs of Rhyniella (three pairs in all) appear to have been similar to each other. Due to relatively poor preservation, the complete morphology of the walking legs remains unknown. It does seem, however, that each leg ended with a distinct single claw. These terminal claws appear to have been relatively long but simple in their structure.
Abdomen
As with modern collembolans Rhyniella possessed six abdominal segments. It appears to have also possessed a furcula ( Whalley & Jarzembowski 1981). In a number of specimens the first abdominal segment has a poorly preserved structure on its ventral surface that may represent the collophore (see insets above).
- Palaeoecology
-
Modern collembolans or springtails are found in a variety of habitats ranging from beaches, caves, grassland, trees and forest floors to the surface of ponds.
They have even been found in Antarctica. In fact unlike many other arthropods it appears collembolans have actually adapted to flourish in relatively cold environments.
On the whole, however, most species seem to prefer cool humid conditions, particularly in soils with an abundance of plant litter. They are mainly saprophagous (living off dead organic, particularly plant, matter) and some species attack young plants. A few modern species are carnivorous feeding on nematodes and other collembolans. In many modern terrestrial ecosystems collembolans play an active role in the breakdown and recycling of dead plant material and, as a result, help in the formation of soils.
It is quite likely that Rhyniella fitted a similar ecological niche within the Rhynie fauna, perhaps living among Rhynia and Aglaophyton stands feeding on soil micro-organisms, spores and breaking down rotting plant litter. This would imply an important interaction between Rhyniella and the Rhynie flora in that it may have helped significantly in the formation of soils during the Early Devonian. It is also interesting that the few specimens of Rhyniella to be discovered appear to be mainly associated with cherts formed within a more aquatic setting (that is, the chert matrix containing the fossils often shows open meshes of filamentous algae, fungal hyphae, cyanobacteria and coprolites). Although it is likely these animals or their moults were washed into localised ponds and areas of standing water, Rhyniella may have been able to live on the surface of ponds buoyed up by surface tension like some modern semi-aquatic collembolan species.
- Other useful references
-
Since Scourfield's time further works have been written relating to Rhyniella.
Among others, useful texts have been written by: Kevan et al. (1975); Rolfe (1980 & 1985); Whalley & Jarzembowski (1981); Crowson (1985); Greenslade & Whalley (1986); Greenslade (1988) and D'Haese (2003).